真菌分类学 Fungi Taxonomy
分类学家的工作是将不同的物种按照它们之间的进化关系分类,以为后续进一步研究和利用这些物种打下坚实的基础。分类学家将一些具有一定相似性的物种归为一类,称为一个物种(简称种),若干个亲缘关系近,演化上分化较晚的物种聚在一起,构成一个属。若干个关系近的属再聚在一起,构成一个科。以此类推,分类的等级从小到大分别是种Species、属Genus、科Family、目Order、纲Class、门Division、界Kingdom,根据这个体系分类学家将不同的物种层层分类。
为了在国际上交流方便,每一个分类群都有其对应的学名。学名是以拉丁文撰写的,一方面拉丁文很少有人使用,语义不会快速变化,另一方面最早的分类体系被林奈提出时便使用了拉丁文,因此这个传统一直保留了下来。
例子Ex 1. 下面是截至2023-07-06,香菇在分类学里的位置(包括每一个分类群及其对应的学名):
真菌界Fungi 双核亚界Dikarya 担子菌门Basidiomycota 蘑菇纲Agaricomycetes 蘑菇亚纲Agaricomycetidae 蘑菇目Agaricales 类脐菇科Omphalotaceae 微香菇属Lentinula 香菇L. edodes
可以注意到,香菇的分类中有“亚界” “亚纲” 这样的阶元。这是因为随着分类学的发展,界门纲目科属种的体系渐渐不够用了,所以新增了亚界、亚门...亚属、亚种等等亚类群。另外,还新提出了很多分类阶元,如“组”是属和种之间的阶元。如果这些体系还不够用,则新增的分类阶元不论大小,统一叫做“分类群(clade)”。
除了种以外,所有分类群的学名都是一个单词。物种的学名由两个单词组成,第一个单词是它对应的属的学名,第二个单词是物种自己的名字,称为种加词。
种加词完全由命名者自己拟定,它可以描述物种的特征,也可以是地名、人名等一些纪念性的名称。属的学名和种的学名在写的时候需要斜体。
通常我们会在学名后面加上命名者的缩写及命名年份,但也可以省略。命名人无需斜体。若某一个物种曾被A学者归入一个属,后来被B学者转移到另一个属,则命名人的格式如此:(A的名称缩写) B的名称缩写 B转移属的年份。
例子Ex. 2
在Ex 1.中,香菇的学名是“L. edodes”。事实上,这是一个缩写,它的完整版本是:
Lentinula edodes (Berk.) Pegler 1976
属名 种名 命名人
若上文已经提到过属名,则下一次写物种的学名时可以缩写属名,如Lentinula edodes缩写为L. edodes。
在这个例子中,Lentinula是微香菇属的学名,也就是香菇所在的属。edodes是种加词,在这里它是“可食用”的意思,描述了香菇的食用性。
(Berk.) Pegler 1976是命名人,这个命名人告诉我们,香菇最早是由Berk.这个人命名的,但当时并未放在这个属,后来在1976年被Pegler转移到微香菇属。
如果你去Index Fungorum上查询具体的信息,可查询到这些缩写对应的学者全名,并了解这个故事的全貌:Miles Joseph Berkeley(缩写Berk.)最早在日本采集到了香菇,并于1878年将香菇命名为Agaricus edodes,这个名称被发表在The Journal of the Linnean Society杂志上,当时置于蘑菇属Agaricus中;后来1976年,David Norman Pegler(缩写Pegler)将这个物种转移到了微香菇属,研究成果发表在Kavaka杂志上。
为了提供一个命名植物、菌物和藻类分类群的稳定方法,国际上制定了《国际藻类、菌物和植物命名法规》(以下简称《法规》),它管理以下这些生物的分类与命名:现存或灭绝的高等植物、藻类(光合原生生物和蓝藻)及其非光合的近亲、真菌(但不含微孢子虫)、地衣、黏菌、卵菌。《法规》每六年更新一次(最新的一版叫做深圳法规),所有的真菌、黏菌等菌物的命名、发表、出现争议时的处理方式都在法规中有明确规定。深圳法规中专门有一章(Chapter F)用于规定被处理为菌物的有机体(含地衣,不含微孢子虫)的名称(在深圳法规发表后,Chapter F又更新了一个版本,即圣胡安版San Juan Chapter F,这一版在英文版深圳法规中缺失,在中文印刷版中存在)。
例如,法规中规定:
为了合格发表,一个处理为菌物的有机体的名称必须:
a) 在1753年5月1日及以后被有效发表(林奈的Species plantarum ed.1处理为在此日期发表)。
b) 除了一些例外 (原文中花了7个条款来阐明这些例外和规定),仅由拉丁文字母组成。
c)符合一些格式上的规定。
譬如:科及以上的分类群处理为复数,科名以-ceae作为后缀,属的名称是主格单数名词,种名由属名和一个形式上为形容词、属格名词或同位词的种加词构,且种加词不可与属名重复等等(原文中,诸如此列的规定有21页)。
(整理自《深圳法规》条款32.1,经过整理,并非法规原文)
《法规》中一些重要的概念
原白protologue
原白是发表一个物种时所有相关联的内容的统称。包括对这个物种的描述、特征集要(diagnosis,概括了这个类群有别于其他类群的特征)、图示、文献、讨论和评论,等等。一个分类群若发表时缺乏描述或特征集要,则它是一个裸名。
模式type
法规中规定,每一个分类群都必须指定一个模式。对于物种,每一个物种需要指定至少一份标本作为模式标本。对于更高级的分类群,需要指定其下级的某一个类群作为模式,例如属需要指定一个种作为模式种,科需要指定一个属或者一个种作为模式,目需要指定一个科、属或种作为模式。
模式是这个分类群的名称所永久依附的成分。例如,鬼伞属Coprinus是一个曾经包含上百个物种的属,它的模式种是毛头鬼伞。后来,系统发育学研究表明鬼伞属应拆成两个枝条,毛头鬼伞及其他零星几个种是蘑菇科的成员,剩余的数百个物种则应该归入另一个科。因为毛头鬼伞是鬼伞属的模式种,所以鬼伞属永远依附毛头鬼伞,被移动到蘑菇科中。这也导致剩余的几百个物种根据法规规定,不得不全部更名。
诸如此类的分类学变动,如果有学者认为可能会造成命名的混乱性(比如这里有几百个物种被更名),可以向国际菌物学大会提议破例处理。2001年,Per M. Jørgensen等人曾提议破例处理,将鬼伞属处理为一个保留名称,留给剩余的数百个物种。不过,此提议没有被通过,理由之一是毛头鬼伞一个物种的知名度要高过那剩余几百个物种的知名度。
物种的模式标本分为多种。
在最早期的时候,分类学家将不会移动,和植物一样生长发育的菌类归入植物当中。这种错误的观点一直持续到1969年五界分类系统的诞生,Whittaker将生物分为原核生物界、原生生物界、植物界、真菌界和动物界,标志着菌物脱离了植物的范畴,独立为界,明确了菌物与其他生物的区别,这一观点成为当时的主流 1(尽管当时真菌界的定义比现在要更广泛,包含了几乎全部菌物)。
比较高等的菌物类群通常难以形成化石,缺乏化石证据来推测演化关系;而比较低等的菌物类群往往形态上相似,物种多而繁杂;加之趋同演化趋异演化、水平基因转移和表观遗传等的影响,使菌物的分类一直谜团重重。二十世纪九十年代以来的系统发育学为此带来了曙光。菌物开始朝多界化发展,各类菌物在演化之树上的地位渐渐明晰。
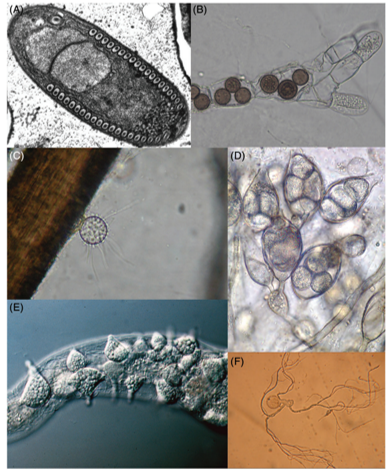
拥有游动孢子的真菌(即传统上的壶菌)。按现在的分类来看,图A为微孢子虫(Fibrillanosema crangonycis),图B为罗兹菌类(Rozella allomycis)孢子囊寄生在芽孢霉门物种(Allomyces sp.)中,图C为狭义壶菌(Rhizophidium keratinophylum),图D为狭义壶菌(Gonapodya polymorpha),图E为芽孢霉门物种(Catenaria anguillulae)寄生在线虫中,图F为新丽鞭毛菌(Neoallimastix frontalis) 图片来源8
真菌界下的高级分类阶元也重新洗牌。在系统发育学研究前,除了纯粹因为完整的生活史不明而被建立的半知菌门外,真菌一般被分为4个门:担子菌门、子囊菌门、壶菌门(水生且孢子有鞭毛)和接合菌门(陆生且产生接合孢子) 12 。2006年,James et al. 10 重建了早期真菌的演化历程,发现传统上的“壶菌”并非单系,其中油壶菌属Olpidium和传统上的接合菌门为姊妹群,Rozella和微孢子虫近缘。2007年,Hibbett等67位菌物学家将真菌界下分为7个门 2 :微孢子虫门Microsporidia、壶菌门Chytridiomycota、新丽鞭毛菌门Neocallimastigomycota、芽孢霉门Blastocladiomycota、球囊菌门Glomeromycota,和双核亚界Dikarya下的担子菌门Basidiomycota和子囊菌门Ascomycota,另还有未定分类的毛霉亚门Mucoromycotina、虫霉亚门Entomophthoromycotina、捕虫霉亚门Zoopagomycotina、与Kickxellomycotina亚门,这一分类奠定了真菌界分类的大致图景。此研究中还证明传统意义上的接合菌门Zygomycota并非单系。此研究中并未将如Rozella等的原始真菌类群放入任何高级分类阶元中。
M.D.M. Jones et al. 2011 3 将罗兹菌属Rozella及近缘的类群建立了隐真菌门Cryptomycota,是菌物学中第一个根据分子数据而非形态特征建立的门(建立此门所基于序列很大一部分来自于环境样本environmental samples,即对土壤、水体等环境采样后直接进行DNA分析,其中测定到很多序列无法找到“真身”,也就无法获得它们的形态特征)。隐真菌门中的序列来自于各种各样的环境,例如土壤、海洋和淡水等环境中,表明这些环境中还有大量未命名和研究的原始真菌物种。Jones et al.认为隐真菌门(此后被称为罗兹菌门Rozellomycota)缺乏真菌的特征,是介于动物和真菌之间的过渡类群,但此后的研究认为罗兹菌门应该是特殊的、退化的真菌类群,它们可能起源于一个非常正常的真菌祖先 13 。罗兹菌门、微孢子虫类Microsporidia及Aphelidea属是真菌界最早分化出的三个单细胞类群。这些类群的生活史很复杂,后来的研究关于这三个类群的分类一直存在争议。罗兹菌门的一些成员在系统学上似乎与微孢子虫关系更近,导致一部分学者将微孢子虫归入罗兹菌门,如Tedersoo et al. 2018 5 ,部分学者则选择拓宽微孢子虫的定义让两个类群都保持单系 12。后文中我们用“罗兹菌+微孢子虫”来指代这一类生物。部分学者认为“罗兹菌+微孢子虫”和Aphelidea并非单系,所以应该归入真菌中,而这三个类群谁先分化出来还需要进一步研究,Tedersoo et al. 2018 5 认为“罗兹菌+微孢子虫”是最早分化出来的类群;也有学者认为两者构成单系群且这个单系群和其他真菌构成姊妹群,他们将这一类群称为后孢菌类Opisthosproidia(或ARM类) 7 8 9 。
Spatafora et al. 2016 4 指出“接合菌门”包含两个主要的类群,且这两个类群并不能构成一个单系群。这两个类群被命名为毛霉门Mucoromycota(包含了Hibbett系统中的毛霉亚门和球囊菌的物种)和捕虫霉门Zoopagomycota(包含了Hibbett系统中的虫霉亚门、捕虫霉亚门和Kickxellomycotina亚门的物种)。大多数研究都支持这一观点 10 11 。Tedersoo et al. 2018基于捕虫霉门下的蛙粪霉属Basidiobolus独特的系统学位置,将其独立为亚界。
Yuanning Li et al. 2019 9 基于真菌基因组重建了真菌的演化之树,他们在构建真菌界的串联树和溯祖树时发现两棵树对后孢菌门的单系性存在争议,这可能是因为后孢菌门中的序列数量太少导致的分歧。但两棵树中都支持罗兹菌门是并系群,微孢子虫应属于罗兹菌。研究中还认为,毛霉门和捕虫霉门能构成一个单系群,故接合菌门可能是单系的,且捕虫霉门的蛙粪霉属与毛霉门构成姊妹群,说明捕虫霉门并非单系。
此分类树基本参照Tedersoo et al. 2018 5 ,Tedersoo et al.的分类树中将许多原本处于亚门的分类阶元提升到了门。另做了以下改动:如上文所述,“后孢菌类”的单系性争议较大,微孢子虫是否要归入罗兹菌也有争议,所以这里将三个类群并列,并统一扔在后孢菌中。蛙粪霉属Basidiobolus在不同研究中的系统学地位有争议 5 8 9 14,故这里采用保守的分类方式,将其算作捕虫霉门的一员。
[1] 李玉,刘淑艳主编. 2015. 菌物学. 北京:科学出版社
[2] D.S. Hibbet et al. 2007. A higher-level phylogenetic classification of the Fungi. Mycological Research, 111(5), 509–547.
[3] Meredith D. M. Jones et al. 2011. Discovery of novel intermediate forms redefines the fungal tree of life. Nature 474(7350): 200-203
[4] J.W. Spatafora et al. 2016. A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data. Mycologia, 108(5), 1028–1046.
[5] Tedersoo et al. 2018. High-level classification of the Fungi and a tool for evolutionary ecological analyses. Fungal Diversity 90(1)
[6] Torruella et al. 2018. Global transcriptome analysis of the aphelid Paraphelidium tribonemae supports the phagotrophic origin of fungi. Communications Biology 1(1)
[7] Karpov et al. 2014. Morphology, phylogeny, and ecology of the aphelids (Aphelidea, Opisthokonta) and proposal for the new superphylum Opisthosporidia. Frontiers in Microbiology 5
[8] Miguel A. Naranjo-Ortiz, Toni Gabaldon. 2019. Fungal evolution: diversity, taxonomy and phylogeny of the Fungi. Biological Reviews 94: 2101–2137
[9] Yuanning Li et al. 2021. A genome-scale phylogeny of the kingdom Fungi. Current Biology 31(8):1653-1665
[10] James et al. 2006. Reconstructing the early evolution of Fungi using a six-gene phylogeny. Nature 443(7113): 818-822
[11] Y. Liu et al. 2009. Phylogenomic analyses predict sistergroup relationship of nucleariids and Fungi and paraphyly of zygomycetes with significant support. BMC Evolutionary Biology 9(1) 272
[12] Kerstin Voigt et al. 2021. Early-diverging fungal phyla: taxonomy, species concept, ecology, distribution, anthropogenic impact, and novel phylogenetic proposals. Fungal Diversity 109: 59–98
[13] T.Y. James, M.L. Berbee. 2012. No jacket required—new fungal lineage defies dress code: recently described zoosporic fungi lack a cell wall during trophic phase. BioEssays 34(2):94–102
[14] Ying Chang et al. 2021.Genome-scale phylogenetic analyses confirm Olpidium as the closest living zoosporic fungus to the non-flagellated, terrestrial fungi. Scientific Reports volume 11
[15] T.Y. James et al. 2020. Toward a Fully Resolved Fungal Tree of Life. Annual Review of Microbiology, 74(1)
[16] Karen L. Haag et al. 2014. Evolution of a morphological novelty occurred before genome compaction in a lineage of extreme parasites. Proceedings of the National Academy of Sciences, 111(43), 15480–15485
[17] Yanji Xu, Louis M. Weiss. 2005. The microsporidian polar tube: A highly specialised invasion organelle. International Journal for Parasitology, 35(9), 941–953
微孢子虫门Microsporidia
微孢子虫是一类极端的寄生虫,以昆虫、脊椎动物(包括鱼类、哺乳动物含人类、鸟类)等为寄主。它们长期坚持寄生主义道路,以至于很多细胞器发生了退化。微孢子虫缺乏真正意义上的,拥有线粒体DNA的线粒体,取而代之的是一个没有DNA的极度退化的细胞器——mitosomes,它参与一些基本的活动如合成铁硫簇,但已经失去了产生ATP的能力。Mitosporidium daphniae是唯一的例外,它们的线粒体中还有DNA。 16 科学家甚至一度怀疑微孢子虫是在真核生物获得线粒体之前就分化出来的一支原始生物,但它们的基因组中还有线粒体相关的基因,且在系统发育树上它们位于真菌的基部位置 8 。它们还退化了鞭毛 ,并魔改了自己的高尔基体,使其成为一种特化的,可以伸出并插入寄主的构造——极管(polar tube)。极管在孢子内是缠绕压缩起来的,当它完全伸出孢子外后其长度看起来壁压缩时更长,宽度也会变宽,形成一个中空的管道来注入感染性的孢子质,这种伸出的机制不明,过程可能类似于将手套的一个指头翻过来。 17 微孢子虫演化出微丝时可能伴随着鞭毛的丢失,两者是重叠的 10 。

微孢子虫孢子的示意图(左)和实拍图(右)。示意图中可见孢子中的极管包含一段竖直部分(manubrium,M)和一段缠绕卷曲的部分(posterior region,PT),极管着生在一个固定盘(anchoring disc,AD)上,竖直部分周围被极质体(polaroplast,在左图中标记为Pl,右图中标记为P)包裹。Nu为细胞核。来源文献17
罗兹菌门Rozellomycota(隐真菌门Cryptomycota、Rozellidea)
罗兹菌门包含了一系列单细胞的内寄生虫,它们拥有游动孢子,寄主为其他拥有游动孢子的真菌、卵菌和一些绿藻。以最常见和广泛研究的R. allomycis为代表,其生活史如下:带有鞭毛的游动孢子具有感染性,会附着在寄主细胞上,之后鞭毛消失,罗兹菌在寄主表面形成一个拥有细胞壁的囊泡,同时它削弱寄主的细胞壁,朝寄主中插入一根管道。原生质会通过管道侵入寄主,生长直到占领所有可利用的空间,随后它在寄主中产生游动孢子囊,释放游动孢子。一些罗兹菌物种可以产生休眠孢子。在寄生和营养阶段,罗兹菌是没有细胞壁的,但它们的休眠孢子拥有细胞壁。培养它们必须与其寄主共同培养。8 13
Jones et al. 2011 3用染色法测试当时能获得的少量样本,细胞壁并未发现几丁质,不符合真菌界的定义,故他认为隐真菌是介于动物和真菌之间的过渡类群;但James & Berbee 2012 13 用同样的方法对未成熟的休眠孢子染色,发现细胞壁中含有几丁质或纤维素,同时对R. allomycis的测序表明它们拥有真菌特有几丁质合成酶,因此他们提出罗兹菌应该是退化的真菌。
James & Berbee 2012 13 出于“合理化分类学命名的精神”,指出一个高等分类阶元的名称应基于其模式类群的学名,罗兹菌门的学名应该是其模式属罗兹菌属Rozella加上代表门的后缀-mycota,即Rozellomycota,Rozellidea不符合此原则,而隐真菌门Cryptomycota所对应的属Cryptomyces已经被用于子囊菌门的一个属,因此这两个名称被废弃。这个学名在2013年被正式发表。
罗兹菌门是一个很多样的类群,依然还有大量待发现的物种,而此前的研究多侧重于可以培养的那些物种。这一类群对我们而言还有许多未知。
油壶菌属Olpidium
上文中仅提到了一下这个属,那么在这里就正式地介绍一下它。油壶菌属是一个包含了约30个物种的小属,基本都是藻类、植物、真菌和小型动物的内寄生虫。它们会产生游动孢子,但系统发育学研究表明它们与陆地真菌是密切相关的,甚至可能是与陆地真菌关系最近的一类,意味着在两者分化后的不久陆地真菌的祖先可能就经历了一次鞭毛退化事件 14 (补充:有研究表明真菌演化过程中至少经历了4次鞭毛退化事件 10 )。它们和罗兹菌的区别是它们生活史各个阶段都有(可能是几丁质的)细胞壁 15 。